
Description
- Appearance: White/Blue with lens
- • Length of strip: 86mm
- • Width of strip: 5mm
- • Length of device: 140mm
- • Width of device: 15mm

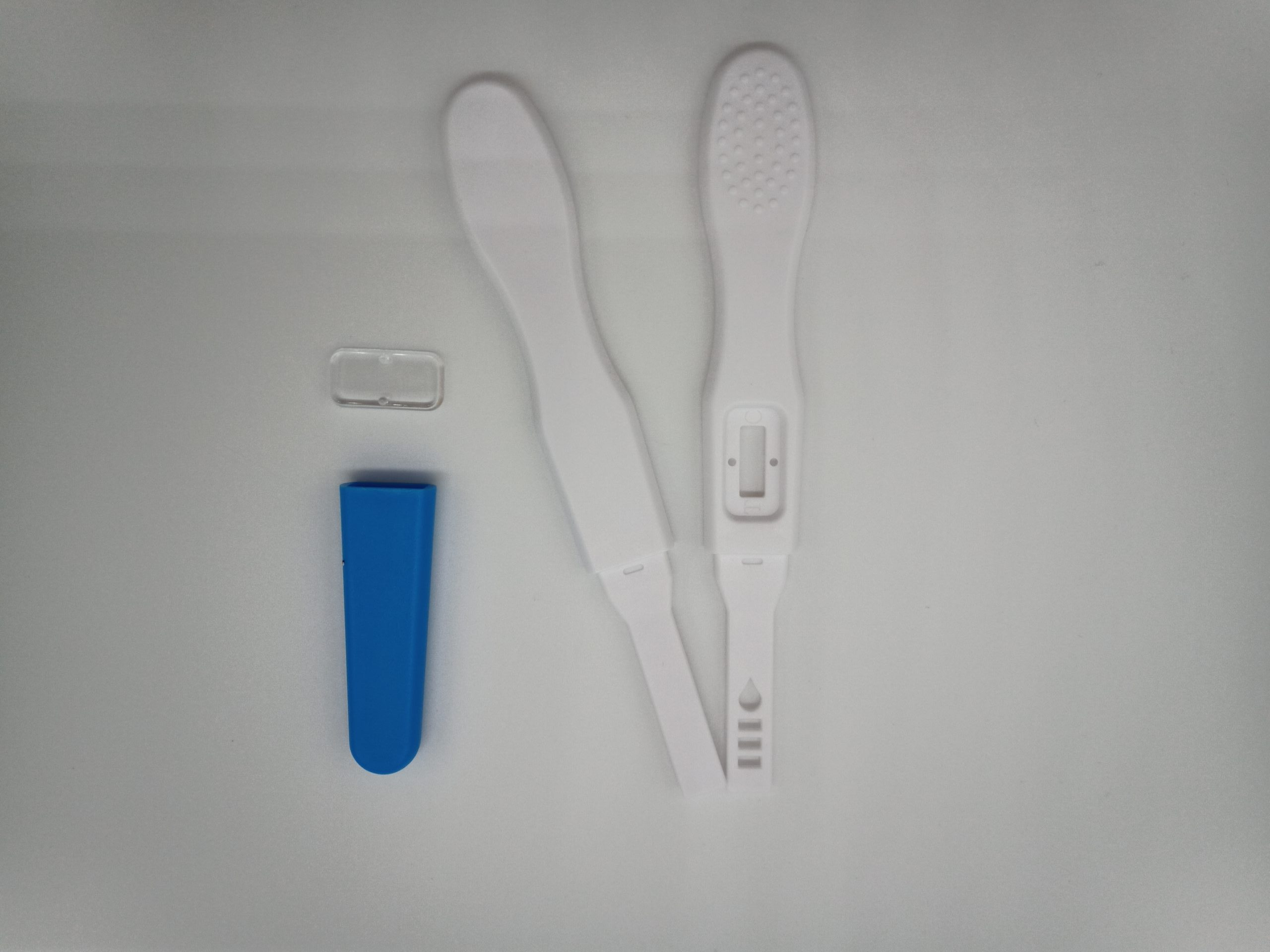

K-LACSU
100 assays of each per kit
| Content: | 100 assays of each per kit |
| Shipping Temperature: | Ambient |
| Storage Temperature: | Short term stability: 2-8oC, Long term stability: See individual component labels |
| Stability: | > 2 years under recommended storage conditions |
| Analyte: | D-Glucose, Lactose, Sucrose |
| Assay Format: | Spectrophotometer |
| Detection Method: | Absorbance |
| Wavelength (nm): | 510 |
| Signal Response: | Increase |
| Limit of Detection: | 100 mg/L |
| Reaction Time (min): | ~ 60 min |
| Application examples: | Flours, beverages, dairy products, milk, foodstuffs containing milk, cosmetics, pharmaceuticals and other materials (e.g. biological cultures, samples, etc.). |
| Method recognition: | Used and accepted in food analysis |
This product has been discontinued (read more).
The Lactose/Sucrose/D-Glucose assay kit is suitable for the measurement and analysis of sucrose, lactose and D-glucose in flour mixtures and other materials.
Explore our full range of mono/disaccharide assay kits.

Advantages
The Lactose/Sucrose/D-Glucose assay kit is suitable for the measurement and analysis of sucrose, lactose and D-glucose in flour mixtures and other materials.

Sulfo DBCO-PEG4-Maleimide is a water-soluble click chemistry reagent with a maleimide group and a DBCO moiety. The hydrophilic PEG spacer arm improves solubility in aqueous buffers. Maleimide group specifically and efficiently reacts with thiols to form stable thioether bonds. The low mass weight will add minimal spacer to modified molecules and will enable simple and efficient incorporation of DBCO moiety onto cysteine-containing peptides or other thiol-containing biomolecules. DBCO is commonly used for copper-free Click Chemistry reactions. Reagent grade, for research purpose. Please contact us for GMP-grade inquiries.

Sulfo DBCO-PEG4-Maleimide is a water-soluble click chemistry reagent with a maleimide group and a DBCO moiety. The hydrophilic PEG spacer arm improves solubility in aqueous buffers. Maleimide group specifically and efficiently reacts with thiols to form stable thioether bonds. The low mass weight will add minimal spacer to modified molecules and will enable simple and efficient incorporation of DBCO moiety onto cysteine-containing peptides or other thiol-containing biomolecules. DBCO is commonly used for copper-free Click Chemistry reactions. Reagent grade, for research purpose. Please contact us for GMP-grade inquiries.

10 Layer Cell Factory Double Narrow Mouth
Huayi Cell Factory is a robust and easy-to-use cell culture platform for applications in the production of human and animal vaccines, therapeutic proteins, cell therapy, and gene therapy.
To address the needs of your workflow, We have three kinds of mouth, which is available on most cell culture products to ensure consistent performance from lot to lot and from format to format.
Specifications: 1 layer 2 layers, 5 layers, 10 layers and 40 layers.
PRODUCT FEATURES
*The product is made of medical grade USP CLASS VI polymer polystyrene
*The product assembled with ultrasonic welded technology
of one 10-layer Cell Factory unit is equivalent to the area
of 36 T-175 flasks

Huayi Cell Factory is a robust and easy-to-use cell culture platform for applications in the production of human and animal vaccines, therapeutic proteins, cell therapy, and gene therapy.
To address the needs of your workflow, We have three kinds of mouth, which is available on most cell culture products to ensure consistent performance from lot to lot and from format to format.
Specifications: 1 layer 2 layers, 5 layers, 10 layers and 40 layers.