

Description
ExcelRT™ Reverse Transcription Kit is a complete, efficient and convenient kit to synthesize high quality first strand cDNA. This kit contains ExcelRT™ Reverse Transcriptase, which is able to synthesize the first strand cDNA at 37~50°C. The ExcelRT™ Reverse Transcriptase is a recombinant Moloney Murine Leukemia Virus (M-MLV) reverse transcriptase, which is designed to reduce RNase H activity and create better thermal stability. This kit also contains RNAok™ RNase Inhibitor, which is active against RNase A, RNase B, and RNase C. This product is supplied with oligo (dT)20 and random hexamers, which are used to synthesize cDNA from poly(A) tailed mRNA and total RNA, respectively.
Features
- Contains all components for reverse transcription
- High yield
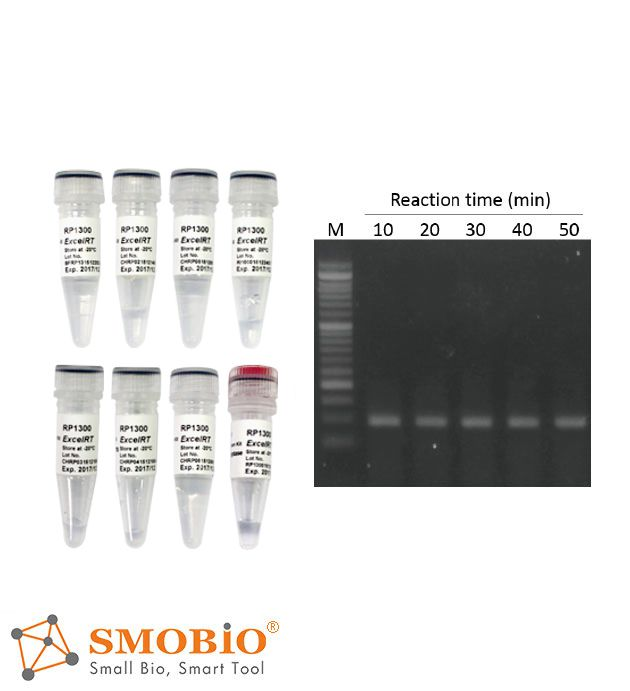
- Thermostable, up to 50°C, during first strand synthesis
- High processivity, generating cDNA up to 8 kb
- Reduced RNase H ribonuclease activity
Application
- Generation of first strand cDNA from total RNA or mRNA.
- Suitable for generating cDNA from RNA with strong secondary structure which can be reduced at temperature up to 50°C.
Storage
-20°C for 24 months

High yield
High sensitivity
Thermostable, up to 50°C, during first strand synthesis
Contents
| ComponentVolume ExcelRT™ Reverse Transcriptase (200 U/μl) 100 μl RNase Inhibitor (20 U/μl)100 μl 5X RT Buffer (DTT)500 μl dNTPs Mix (10 mM each)200 μl Oligo (dT)20 (50 μM)100 μl Random Hexamers (100 μM)100 μl DEPC-Treated H2O 1 ml x 2 |
Storage Buffer
Reverse Transcriptase: 20 mM Tris-HCl (pH 7.5), 200 mM NaCl, 0.1 mM EDTA, 1 mM DTT, stabilizer and 50% (v/v) glycerol
RNase Inhibitor: 40 mM HEPES-KOH (pH 7.5), 100 mM KCl, 8 mM DTT, 0.1 mM EDTA, stabilizer and 50% (v/v) glycerol
5X RT buffer (DTT)
250 mM Tris-HCl (pH 8.3), 375 mM KCl, 15 mM MgCl2 and 50 mM DTT
Storage
-20°C for 24 months



